John A. Medeiros
![]()
The achromatic receptors of night vision (scotopic vision) are the rods, so-named because of their shape. The color vision receptors of daylight vision (photopic vision) are the cones, also so-named because of their shape. The duplicity theory of the eye in terms of the operation of these two receptor types has been known for a long time, but there has never been any explanation for the universal dichotomy of receptor structure. That dichotomy is apparent in the classic rod and cone drawing by Schultze (1866) who first clearly articulated the duplicity theory of the retina (see figure below).
Surely nature didn’t build the two receptor types with different shapes just so researchers could tell them apart!
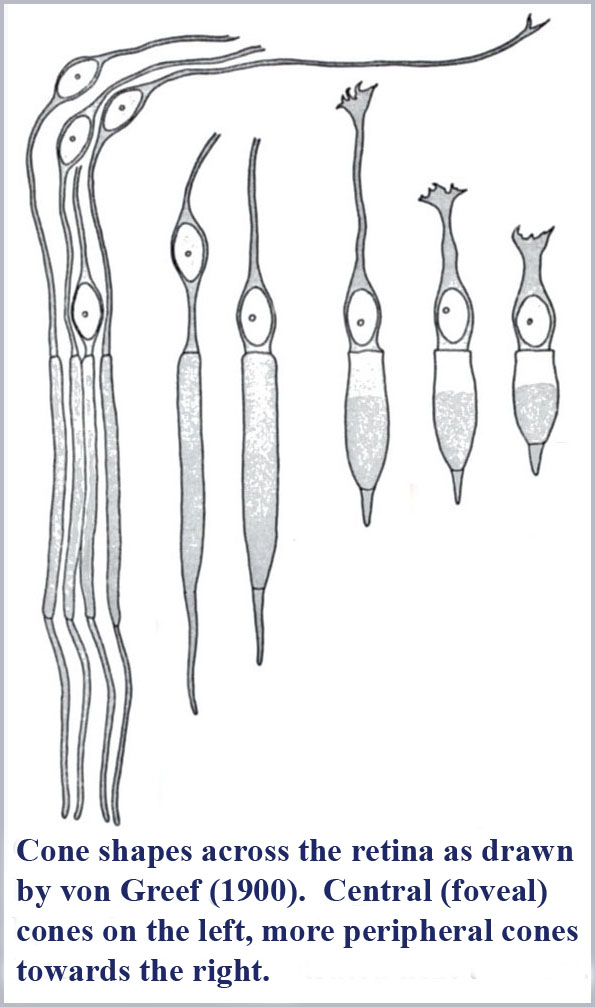
It is also notable that the cone shape is not identical throughout the human retina. The photosensitive outer segment portion of the cones varies systematically from being long and gently tapering in the central (foveal) part of the retina to being shorter and squatter (more obviously tapering) in the peripheral portions of the retina (von Greef's drawings of the cone morphology over various regions of the retina are shown below). Coincidently, the color vision function provided by the cones also varies systematically from the center to the peripheral portions of the retina. Color vision has the best resolution (three-dimensional or trichromatic) where the cones are long and gently tapering, decreases in resolution (to be essentially two-dimensional or dichromatic) in intermediate areas of the retina, and decreases further to its lowest resolution (essentially one-dimensional or monochromatic) in the far periphery of the retina (although color vision function is not totally lost for sufficiently bright or large patches of color).
So, why is there a systematic shape change of the cones across the retina and why is it apparently correlated with color vision function? Is this just coincidental?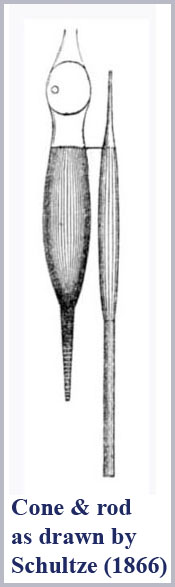
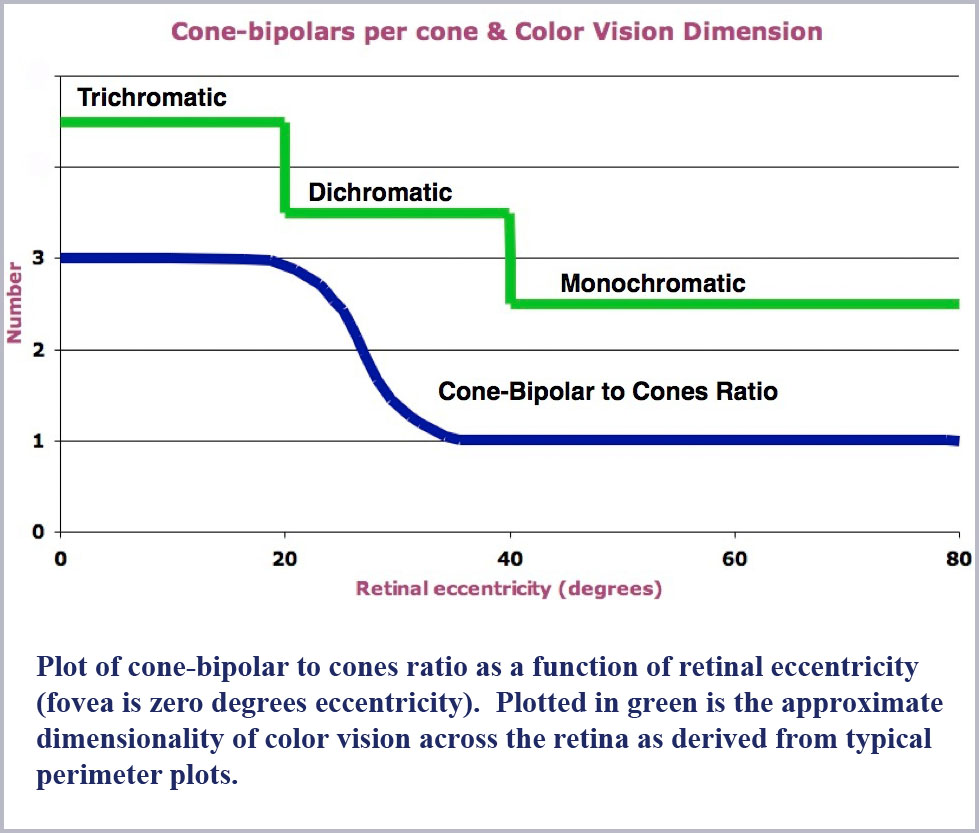
The signals generated by the retinal cones are relayed through the bipolar cells with which they make synaptic contact. (The bipolars in turn contact the ganglion cells which relay the visual information on to the brain.) Cell counting studies (Missotten, 1974; Vilter, 1949; c.f. Sheppard, 1968) have shown that the number of bipolars per cone is correlated with the dimensionality of the color vision provided in each area of the retina. That is, there are three bipolars for each cone where color vision is trichromatic in the center of the retina, two bipolars per cone in the intermediate areas where color vision is dichromatic and but one bipolar per cone where color vision is monochromatic in the retinal periphery (see the figure below). Is this too a coincidence?
In any case, besides having a conical shape, the color receptors are small, with the diameter of their photosensitive outer segments barely larger than the wavelength of light itself. That the photosensitive portion of the cones are so much smaller than the minimum resolvable spot of light focused on the retina by the eye's optics and that they are tapered make no sense in the standard three-cone model of color vision. However, these two bits of information are critical jumping off points to describe the physics of what is happening to light in these receptors and perhaps a new understanding of human color vision.
If one tries to model how light propagates within the cones, one finds that you have to employ waveguide mode physics computations. The details of this computation may be complicated, but the physical processes that are going on are not.